Das Herz
Eine Druck-Saugpumpe mit einem ausgeklügelten ökonomischen Energierückgewinnungssystem
Das Herz leistet zwischen 90.000 und 120.000 Kontraktionen pro Tag. Je nach körperlicher Belastung und psychischen Stress kann es sich jeder Lebenslage anpassen, ohne dabei seine Mikrostruktur zu verändern. Es transportiert ca. 7500 Liter Blut pro Tag und schlägt bis zum 75. Lebensjahr eines Menschen mehr als 2 Milliarden mal. Nach dem Maximum Prinzip aus der Ökonomie heißt das, dass das Herz mit seinen gegebenen Mitteln eine maximale Leistung erbringen kann und dabei noch Energie von bis zu 25 % seines Energieaufwandes einspart. Wie könnte man sich ein lebendiges Herz sonst vorstellen, wenn es nicht durch Energie in Schwung gehalten wird und durch Informationen gesteuert wird wie intensiv und wie oft es schlagen soll. Man betrachtet es für den Organismus als Verwaltung zur Wiederherstellung und Erhaltung der Homöostase. Hier stellt sich nun die Frage nach der Gültigkeit des Frank-Starling-Gesetzes aus dem 19. Jahrhundert. 1988 haben T.F. Robinson, S. M. Factor und E. Sonnenblick nämlich ein neues Modell vorgestellt.
Dieses besagt, dass eine dynamische Beziehung zwischen Systole und Diastole entscheidend für die Funktion des Herzens ist. Sie beschreiben in ihrem Modell eine Wechselbeziehung zwischen potentieller Energie und kinetischer Energie die während einer Herzaktion entsteht. Ähnlich dem Prinzip moderner Schnellfeuerwaffen, bei denen die Kraft eines Feuerstoßes die Nachladungsenergie für die nächste Salve liefert. Auf das Herzen bezogen heißt das, dass die Energie die bei jeder Systole entsteht, gespeichert wird und in der darauffolgenden Diastole wirksam wird. Damit ist gemeint, dass die potentiell gespeicherte Energie in eine kinetische Energie und die kinetische Energie wiederum in potentielle Energie umgewandelt wird. Ob diese neue, andere Denkweise wirklich eine Reformation des Frank-Starling-Modells nötig macht, wird im folgenden Artikel versucht näher zu erörtern.
Mechanismus des neuen Modells
Die Kontraktion der beiden Ventrikel des Herzens treibt die Blutsäule nach superior in die abführenden Arterien und das Herz selbst wird durch die Rückstoßkräfte in kaudaler Richtung bewegt. Hierbei kommt es zu einer Dehnung der großen Blutgefäße wie Aorta, der Arteria Carotis communis, Arteria subclavia und den bindegewebigen Mikrostrukturen des Herzens. Die Kräfte, die dagegen vom Ventrikelmyokard aufgebracht werden, wobei die Wandspannung der Pulmonalarterie und Aorta überwunden wird, heißt Nachlast. Durch den so entstehenden Unterdruck und den Venendruck der herznahen Zentralvenen wird Blut angesaugt und die Vorhöfe vergrößern sich dadurch. Das Herz macht hier eine Bewegung nach superior was die Füllung der Vorhöfe unterstützt. Die Kraft, die am Ende der Diastole zu einer Dehnung der kontraktilen Herzmuskelfasern führt nennt man Vorlast. Bis zu diesem Punkt hat das Frank-Starling-Gesetz noch seine Gültigkeit. Doch das neue Modell von T.F. Robinson et al. sagt aus, dass während der Füllung der beiden Vorhöfe wieder Energie für die nächste Systole aufgenommen wird. Es ist ein Selbstregulierungsmechanismus. Ein Wechselspiel zwischen der Umwandlung von potenzieller Energie zu kinetischer Energie und umgekehrt. Form, Funktion, Struktur und Bewegung sind untrennbar mit einander verbunden. Wie in Abbildung I kann man sehen, wie sich die Energie auf Grund der Faserrichtung von der Mikrofaserrichtung bis in die Makrofaserrichtung fortsetzen kann.
Während der Herzaktion in der Systole macht das Herz eine Torsionsbewegung gegen Uhrzeigersinn und während der Diastole eine im Uhrzeigersinn. Ein Teil der kinetischen Energie wird während der Systole gespeichert, um die Kammern während der Diastole zu füllen. Ähnlich wie das Prinzip eines Punchingballs, wo ein freibeweglicher Boxball an einem Aufhängesystem befestigt ist. Bei jedem Schlag mit der Faust gegen den Ball werden große Energien freigesetzt und gespeichert. Übertragen auf das Prinzip des Herzens mit seiner Vor – und Nachlast ist die Makrostruktur mit seiner faszialen Relation zu seiner Umgebung, (in unserem Modell ist hier das Aufhängesystem des Punchingballs) und die Mikrostruktur, (die Beschaffenheit der Federzügel und der Boxballs) die die Fähigkeit haben, potentielle Energie zu speichern und wieder in kinetischer Energie in Form von Bewegung umzuwandeln und abzugeben.

In dieser Abbildung kann man die Bewegungswelle, die sich von der Mikro- bis in die Makrostruktur weiterleitet. In Braun gekennzeichnet die Relationen des Perikards. In Rot die Torsionsbewegung des Herzen während einer Herzaktion . In Blau gezeichneten Pfeile die weiterlaufenden Bewegungen im faszialen Bereich.
Die unterschiedlichen Strukturen des Herzens und ihre Bedeutung
Das Herz befindet sich im Mediastinum medius in der inferior gelegenen Regio Cardiaca. Hier sind die wichtigsten Relationen des Perikards zu seiner Umgebung.
Zum größten Teil, ist das Herz in den elastischen Lungenteilen als weiches Kissen rechts und links eingebettet. (Abb.II) Das Perikard wird von lockerem Bindegewebe umhüllt welches sich von der Pleura mediastinalis trennt. Die unter dem Herzen liegende Diaphragmakuppel trennt die im Bauchraum oben liegenden Organe vom thorakalen Zylinder. Eine gute Dynamik des Diaphragmas unterstützt die Herzfunktion entscheidend. Die Bewegung, die bei Inspiration und Exspiration entsteht, unterstützt das Herz in seiner Füllungs– und Austreibungsphase (Roemheld 1871-1938).

Abb. H.K. Corning – Lehrbuch der topographischen Anatomie 3. Auflage Auflage 1911.
Die ligamentären Verbindungen des Perikards
In der Recherche nach den Verbindungen des Perikards zu seiner Umgebung ist in den meisten Anatomiebüchern der Neuzeit keine wirklich ausführliche Beschreibung der Verbindungen zurück zu finden. Die beiden französischen Autoren J. Brizon und J. Castian zeigen dagegen in ihrem „ Les Feuilletes D`Anatomie Fascicule 14. (siehe Abb. III)“ eine zumindest deutliche Übersicht. Hieraus lassen sich die Relationen des Perikards mit seinen elastischen Verankerungen zu seiner Umgebung ableiten. Es sind wichtige Makrostrukturen und bieten die Voraussetzung der perikardialen maximalen Mobilität und auch Stabilität in seiner Lage im Brustkorb.
Folgende Strukturen sind zu unterscheiden:
- Lig. Sternopericardiaca
- Lig. Phrenopericardiaca
- Lig. Vertebropericardiaca
- Lig. Costopericardiaca
- Membrana bronchopericardiaca

Abb. J. Brizon und J Castian – Les Feuilletes D`Anatomie Fascicule, rot: Lig. phrenopericardiaca, grün: Lig. Sternopericardiaca, blau: Lig. vertebropericardiaca, braun: longitudinale Fascien posterior Fascia prevertebralis in Kontinuität zum Peritoneum Patieltale posterius und anterius Fascia cervicalis media , Lamina pretrachealis in Kontinuität zum Peritoneum Partietale anterius.
Hinsichtlich der anatomischen Realität dieser Verbindungen sind zwei sehr interessante DO-Arbeiten zu erwähnen. Jürgen Seidler hat in einer D.O. Arbeit von 2004 über das Lig. Vertebropericardiacum bei 15 anatomischen Präparaten lediglich nur an zwei Präparaten eine Verbindung zur Halswirbelsäule gefunden. Die Arbeit von 2006 der beiden Kolleginnen Bartmer-Leitl & Kaufer stellten an Hand von Präparaten fest, dass z.B. beim Lig. Sternopericardiacum kein standardisierter Verlauf des Ligamentes zu beschreiben ist. Sieben von neun Präparaten zeigte deutliche Züge in drei Verlaufsrichtungen auf. Beim Lig Phrenopericardiacum wird in der Literatur eine Verbindung zwischen dem Centrum tendineum und dem Perikard beschrieben. An den von ihnen untersuchten Präparaten zeigte sich nur eine sehr feste, verwachsene Struktur. Im Großen und Ganzen ist das Perikard mit seinen Befestigungen im fazialen System als eine „Zentralsehne“ zu sehen, die sich vom Cranium bis zum kleinen Becken in kranio-kaudaler Richtung erstreckt. An der dorsalen Seite vor der Wirbelsäule liegend, befindet sich die Fascia Prevertebralis die eine Kontinuität mit der Fascia Endothoracica hat. In ihrem weiteren Verlauf nach caudal wieder eine Relation mit dem Peritoneum parietale posterior bildet. Auf der anterioren Seite ist die Fascia cervicalis media von C7 und Os Hyoideum kommen wird im oberen Abschnitt auf Höhe des Hyoids Fascia pretrachealis genannt. Sie verläuft nach caudal und bildet unterhalb des Diaphragmas eine Relation mit dem Peritoneum Parietale anterior. Die diversen Befestigungen des Perikards haben in den jeweiligen Abschnitten eine Relation zu den o.g. Faszien im Mediastinum.
Das Perikard
Die Umhüllung des Herzens besteht aus dem Pericardium fibrosum, dem parietalen Blatt und dem Pericardium serosum, dem viszeralen Blatt. Das Pericardium fibrosum besteht aus sehr derben Kollagenfasern und elastischen Fasernetzen, dessen Züge in einer dreischichtigen, scherengitterartigen Anordnung strukturiert ist. Zudem verknüpft das Pericardium fibrosum den Herzbeutel mit seinen faszialen und ligamentären Relationen dem Aufhängesystem. Das Pericardium serosum besteht aus einschichtigen Plattenepithel und hat ebenfalls zwei Blätter. Zwischen den beiden besteht ein Spaltraum. Dieser ist mit ca. 30 ml der durch das Epikard sezernierten Flüssigkeit, dem Liquor pericardii gefüllt. Diese Flüssigkeit gewährleistet während der Herzaktion die reibungsarme und freie Verschieblichkeit der beiden Blätter. Das Perikard hat auf Grund seiner Makro –und Mikrostruktur die wichtige Funktion eines Synchronisators der zwei Kräfte die während der Systole entstehen, so aufeinander abstimmt, dass ein Gleichgewicht zwischen der Kraftabsorption von außen und innen entsteht.
Die embryologische Entwicklung
Hinsichtlich der embryologischen Entwicklung ist es interessant zu erkennen, dass die komplexe Entwicklung und die daraus resultierende Form des Herzens mit dem des neuen Modells entsprechen. Mehr noch, die gesamte Entwicklungsbewegung wird von Francisco Torrent-Guasp (1935 – 2005) in der Form des sog. „helical ventricular myocardial band“ näher umschrieben.

Abb. von F. Torrent-Guasp Hearthelix „helical ventricular myocardial band“ Die Bewegung während der Füllung und Austreibungsphase. Rechtes Bild die embryologische Entwicklung in der 3. Woche: Durch Druck –und Torsionskräfte entstehen zuerst zwei Endokardschläuche, die sich durch die Fusion und Rotation um eine Transversalachse sich Atrien, Ventrikel und Septen umbilden. Moore K.L The development human 1988
Die Herzwand
Die Herzwand besteht aus drei Schichten, dem Epikard, Myokard sowie Endokard. Das Epikard liegt außen, es ist ein Blatt des Perikards und ist mit dem Myokard verwachsen. Das Epikard ist identisch mit dem Pericardium serosum. Das Endokard bedeckt die Hohlräume des Herzens und überzieht die Papillarmuskeln. Die Duplikaturen des Endokards bilden die Herzklappen. Es besteht aus einer einschichtigen Lage mit Endothelzellen und einer Basalmembran. Hinsichtlich des Modells spielt das Myokard eine besonders interessante Rolle. Das Myokard ist quergestreift und unterscheidet sich deutlich durch eine Vielzahl von Besonderheiten von der Skelettmuskulatur. Die Arbeitsmuskulatur des Herzens zeigt ebenfalls einen regelmäßigen Aufbau mit einer speziellen Querstreifung der Muskelfasern auf. Das Myokard ist eingebettet zwischen Epikard und Endokard.
Die einzelnen Muskelschichten haben einen äußeren schrägen, mittleren ringförmigen und inneren längsförmigen Verlauf. Präparatorisch lässt sich ein schraubenartiger Verlauf des Myokards feststellen. Von einer äußeren Schicht, die die Kammern umgibt ausgehend, scheren Muskelbündel aus und umfassen in zirkulärer Verlaufsrichtung jede Herzhälfte getrennt. Daher kommt es bei jeder Herzaktion zu einer ventrikulären Torsionsbewegung. Ab jetzt wird es interessant, weil nun das Frank-Starling-Modell Platz räumen muss für das neue Modell. Die vorhergenannten Strukturen sind lediglich das struturelle Fundament. Ein Teil der Kontraktionsenergie muss in der Zelle gespeichert sein, doch wie? Sowohl die Struktur der Sarkomere selbst, als auch Anteile des Zytoskeletts, welches das Zellskelett bildet und damit die Form der Zelle aufrecht erhält, müssen laut Robinson und seine Kollegen dafür verantwortlich sein!
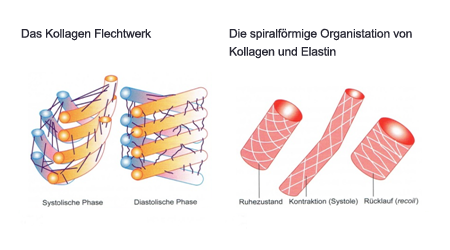
Abb. Verhalten der Bindegewebszügel während der Systole und Diastole
Wenn sich das Sarkomer kontrahiert, treffen sich die Aktinfäden gegenüberliegender Bündel im Zentrum des Sarkomers. Geht dieser Vorgang noch weiter, schieben sich die Aktinfäden innerhalb des Myosingitters übereinander. Durch diese doppelte Überlappung nimmt das zylinderförmige Sarkomer an Umfang zu und zugleich werden die Glanzstreifen und die anderen seitlichen Strukturelemente der Zelle gedehnt. Hier gibt es einen Mechanismus, der einen Teil der Kontraktionsenergie speichert und damit zum anschließenden Strecken der Herzmuskelzelle beiträgt. Auch das Bindegewebe der Muskelzellen speichert zum Teil die Kontraktionsenergie während der Systole. Die Oberfläche der einzelnen Muskelzellen wird von spiralig angeordneten Bindegewebefasern umgeben, die sich mit und gegen dem Uhrzeigersystem anordnen. Diese gekreuzten Faserzüge aus dehnungsstabilen Kollagen mit verschiedenen Anteilen von Elastin und extrazellulären Protein, weisen eine ähnliche Eigenschaft wie Gummi auf. Das elastische Bindegewebe kann sich bis auf 170 Prozent seiner ursprünglichen Länge dehnen. Gleichzeitig schützt das Kollagenfasernetz auch die Sarkomere vor Überdehnung. Es wird vermutet, dass die Netzstruktur des Bindegewebes einen Teil der Rückstellkraft, die bei der Rückdehnung der Muskelzellen zu ihrer ursprünglichen Länge entsteht, speichert. Hier verdickt sich während der Kontraktion die Muskelzelle und drückt gegen das umliegende Gewebe. Nach der Kontraktion pressen die Fasern die Zelle wieder zusammen und verlängern sie. Man kann sich diesen Mechanismus mit dem Prinzip einer Hängematte vorstellen. Die verknüpften Fasern sind relativ fest und starr. Doch trotz der starren Struktur haben diese aufgrund der sich überkreuzten Sisalstränge, ähnlich eines Parallelogramms, die Fähigkeit sich zu überdehnen ohne einen Schaden zu nehmen. Laut Vermutung vom T.F. Robinson et al. trägt das Bindegewebe auch noch zum Entstehen eines diastolischen Soges und somit zur Füllungsphase des Herzens durch die aktive Dehnung bei. Dies erklärt sich durch die seitlichen Verbindungen der Glanzstreifen mit den einzelnen Muskelzellen. Diese Verbindungen sind kordelartig gedreht, was sie sehr zugfest machen, ähnlich dem Prinzip eines Tragseiles einer Hängebrücke. So kann es sein, dass die Glanzstreifen einer Verformung des Herzen entgegenwirken, indem sie die Zellen zusammen binden, und einen Teil der dabei aufgenommenen Energie für den diastolischen Sog zurückhalten. M.a.W. tragen die Glanzstreifen zum Saugeffekt bei, indem sie die Aktivität der einzelnen Muskelzellen koordinieren. Die Spannung in den Glanzstreifen am völlig zusammengezogen Herzen könnte das Speichern von Energie aus der systolischen Kontraktion unterstützen, also wenn es über sein Indifferenzvolumen hinaus kontrahiert.
Fazit und Schlussfolgerung
Zusammenfassend kann man sagen, dass unter der Betrachtung der Mikro-und Makrostrukturen des Herzens, das Frank- Starling- Modell nur zum Teil Gültigkeit hat und sich wirklich nur auf ein insuffizientes Herz beziehen kann. Schaut man sich Form, Struktur und Funktion in seiner Komplexität an, so beeinflussen sie sich gegenseitig und es entsteht ein immer wiederkehrender Mechanismus, nämlich die permanente Umwandlung der potentiellen Energie zu kinetischer Energie und kinetischer Energie in potentieller Energie. Das Modell von Frank-Starling funktioniert unter der Betrachtung der Makrostrukturen und begrenzt die Herzaktion nur auf seiner Vorlast, die Füllung der Herzhohlräume, der enddiastolische Wandspannung. Die Reformation des Gesetzes beruht auf die Betrachtung der Struktur, Form und Funktion seiner Mikrostrukturen. Sie bietet eine Erklärung der Nachlast des Herzens, den Auswurf und das Überwinden der Widerstände in den Gefäßen. Das Modell zeigt ebenfalls wie hierbei gleichzeitig wieder Energie für die nächste Herzaktion zurück gewonnen wird. Jede Struktur des Herzens sei es sein Aufhängesystem, das Perikard, das Myokard bis zu seiner Mikrostruktur die Myokardzelle, haben die Fähigkeit den Wandel zwischen Potentieller Energie und kinetischer Energie zu vollziehen. Ähnlich nach dem Modell einer Automatikuhr, dessen Unruhe sich durch ein Schwungrad und einer Spiralfeder die reibungsfrei gelagert ist, immer wieder Energie aufnehmen kann ohne Energie von einer Batterie zu beziehen. Nur durch die eigene Bewegung des Armes als Impulsgeber seines Uhrenträgers, die freigewordene Energie zu speichern die durch die Bewegung frei gesetzt wird, um kontinuierlich eine genaue Zeigerbewegung zu vollziehen.
Dr. Still sprach immer von einer vollkommenen Form des Herzens bis zu den Arterien und dass die Arbeit des Herzens vollkommen ist, wenn das Blut in die Arterien pumpt (Das große Still Kompendium 2005). Das Herz ist eine Ausdauerpumpe mit einer enormen Energierückgewinnungseigenschaft – ein faszinierendes Organ. In der Tatsache, dass wir durch die Betrachtung unseres Modells auf Grund der Lage und Formen der Makro- und Mikrostrukturen, während unserer Arbeit, sei es im parietalen, viszeralen oder kraniosacralen Bereich, immer wieder Einfluss auf die Funktion des Herzens nehmen können.
Literatur:
- Bartmer-Leitl, Kaufer D.O. Arbeit von 2006 Topografische Untersuchung der Anheftung des Perikardes ( im Mediastinum) zu seinen kaudalen, anterioren und lateralen Nachbarstrukturen
- J. Brizon und J Castian – Les Feuilletes D`Anatomie Fascicule 14
- H.K. Corning – Lehrbuch der topographischen Anatomie 3. Auflage 1911
- Dr. C. Heitzmann – Descriptive und Topographische Anatomie des Menschen 1873
- Horn – Biochemie des Menschen 5. Auflage 2012
- van der Heede Patrik J.L. Danjon – das kardiovaskuläre System in der Osteopathie 2012
- Moore, K.L. – The development human – 1988
- Schiebler, Zilles, Schmidt – Anatomie 8. Auflage 1999
- Seidel Jürgen D.O. Arbeit von 2004 Lig. Vertebropericardiacum
- Spektrum der Wissenschaft – T.F. Robinson, S.M. Factor, E. Sonnenblick
- Still A.T. – Das große Still-Kompendium 2005
- Eigene Aufzeichnungen aus dem Thorax Unterricht Steffi Geldschläger College Sutherland Ulm
- Eigene Aufzeichnungen aus dem Physiologieunterricht von Max Giradin und Mario Kraml College Sutherland